At a time of dramatic worldwide coral reef decline, much of modern coral reef science has necessarily focused on what contributes to the demise of coral reef systems. But for reef conservation and protection efforts to become significantly more effective, it is increasingly important to focus on what actually makes a reef thrive. The aim of this project is to provide and share data on the physical, biological, and ecosystem dynamics that contribute to persistence of endangered coral species, and to place those data in the context of recent geologic time.

Endangered Acropora cervicornis at Coral Gardens, Belize
What: This nine-student Keck Geology project is aimed at assessing the status of living acroporid – dominated coral reefs off Ambergris Caye, Belize. These reefs are unique in the Caribbean region where acroporid-dominated reefs are relatively rare. The goals of this project will be to determine 1) the timing of acroporid dominance of this patch reef system (and whether coral growth has been continuous, intermittent, or re-established after a haitus), 2) whether carbonate accretion at Coral Gardens is in a phase of net growth or decline, 3) the degree of ecological, biological, and physical heterogeneity across this patch reef and potential contributions of heterogeneity to overall community structure, and 4) why these reefs are flourishing against a backdrop of global coral deterioration. This project will use photographic, video, in situ, and laboratory measurements of coral and algal growth, coral morphology, bioeroder and encruster density, water and coral chemistry, and core data to characterize reef ecosystem dynamics in time and space. This project is designed to document, define, and explore the conditions favorable to acroporids at a site of exceptional growth of these endangered coral species.
When: NEW DATES! June 23- July 21, 2014
Where: Ambergris Caye Belize and Lexington, Virginia (Washington and Lee University)
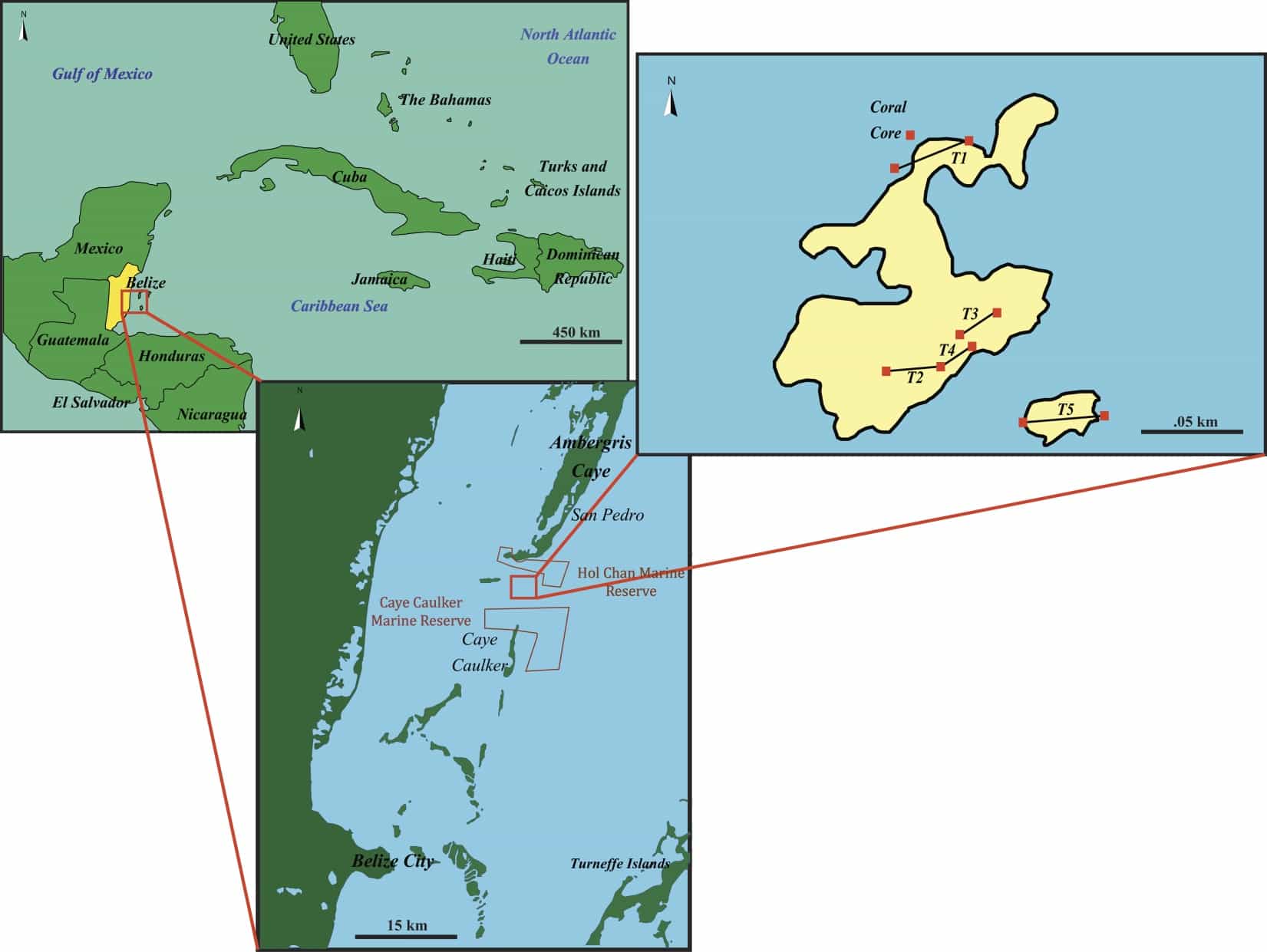
Map of Coral Gardens on the Mesoamerican Barrier Reef off the coast of Belize. Map A shows the relative location in the Caribbean and map B shows the location of Coral Gardens and marine reserves near the study site. Red squares at Coral Gardens (C) mark buoys and straight black lines mark coral transects within the study area.
Who: 9 Keck students and Lisa Greer (Washington and Lee University), Halard Lescinsky (Otterbein University), and Karl Wirth (Macalester College).
Project Overview and Goals
Acropora cervicornis (Staghorn coral) and Acropora palmata (Elkhorn Coral) have been important reef builders throughout the Pleistocene and Holocene (Jackson, 1992; Hubbard et al., 1994: Greenstein et al., 1998). But thriving acroporid populations are now exceptionally rare in the Caribbean and Atlantic (e.g. Precht and Aronson, 2004). Causes sited for the dramatic decline of this (and other coral) species vary, but are virtually all tied directly or indirectly to human-induced environmental or climatic change (e.g. Jackson, 2001; Precht and Aronson, 2004). Yet we still know very little about the specific environmental conditions in which A cervicornis and other acroporid species actually thrive. And there is still some debate about whether or not the current 30+ year absence of Caribbean acroporids is unprecedented with respect to geologic time.



All three acroporid species are found in abundance at Coral Gardens (A: Acropora palmata, B: Acropora prolifera, C: Acropora cervicornis along Transects 1-5).
Acroporid reefs in decline: Acroporid coral species are currently experiencing massive die-offs throughout the Atlantic basin and A. cervicornis and A. palmata are listed as threatened on the U.S. Department of the Interior Endangered and Threatened Wildlife list. Many scientists fear that A cervicornis may be particularly sensitive to environmental change and the demise of the species may be a sign of impending doom for Caribbean reefs in general (e.g. Precht and Aronson, 2004). A primary cause for collapse is White Band Disease (WBD; Rogers, 1985; Aronson and Precht, 2001), but many anthropogenic factors may be enhancing or driving disease effectiveness. Of the human-influenced threats to corals, macroalgae abundance due to overfishing and eutrophication, and climate change are important. In the last few decades reefs have experienced dramatic shifts from coral- to algae-dominated ecosystems (Hughes, 1994; Pandolfi et al., 2005).
The question of whether the recent die-off of acroporids is anomalous with respect to the geologic record has been a subject of debate. Several studies have suggested that there is no evidence of similar phase shifts in acroporid-dominated reefs of the Holocene or Pleistocene. A phase shift might be seen as a hiatus in coral growth or a change in species dominance from Acropora to another coral. Hubbard et al (1994) found long periods of uninterrupted Holocene A. palmata growth in middle to late Holocene cores off St Croix and Aronson and Precht (1997) found similar persistence of late Holocene A. cervicornis reefs in cores from Channel Cay, Belize. Wapnick et al (2004) showed ~800 years of continuous late Holocene A. cervicornis growth in Jamaica with one possible hiatus. Greer et al. (2009) showed 2000 years of continuous A. cervicornis growth in the early Holocene Dominican Republic. These studies suggest that current decline is anomalous and most hypothesize that the current die-off is likely related to anthropogenic environmental or climatic change in conjunction with a well-documented Caribbean-wide outbreak of WBD (Rogers, 1985: Aronson and Precht, 2001).
Most living A. cervicornis today exist in small patches and isolated colonies, and true A. cervicornis-dominated ‘reefs’ are now rare (Miller et al., 2009). As part of this study, we propose to examine whether the acroporid coral growth at Coral Gardens has been continuous over the last several decades (a true refugia) or whether acroporid growth is returning after collapse. Whether this is a site of relatively unique resilience or a signal of coral recovery, we aim to determine what factors are contributing to the unusual health of this reef. If permission is granted by the Belize Fisheries Department, we plan to collect several cores through, or in very close proximity to living coral. Taphonomic and age data will be used to assess how continuous growth was at that location.
 Field Site: This project will primarily take place at a site called Coral Gardens (or Mitchell Rocks), off the southern tip of Ambergris Caye, Belize . Coral Gardens sits between the 1,116 hectare Hol Chan Marine Reserve, created in 1987, and Caye Caulker Marine Protected Areas, but Coral Gardens has no protected status. Hol Chan is the oldest marine reserve in Belize and ‘no-take’ regulations are actively enforced within the park. The heart of Coral Gardens is composed of acroporid-dominated patches that are variably connected to one another. Five semi-permanent transect locations were established in 2012 and each transect end has been marked with rebar stakes, underwater buoys, and high resolution GPS measurements. Preliminary data from three field studies at Coral Gardens in 2011, 2012, and 2013, suggest that A. cervicornis populations (as well as A. palmata and the hybrid species A. prolifera) are anomalously healthy and robust at Coral Gardens, Belize. All three acroporid species (A. cervicornis, A. palmata, and A. prolifera) are present, live acroporid tissue abundance is high in many places, actively growing branch tips are common, and new coral recruits can be seen colonizing recently dead coral rubble and framework. Why is this isolated patch “healthier” than the surrounding acroporid-dominated patches? How do grazer dynamics, hydrology, and other physical and biological variables differ between sites? How might these different reefs look in the geologic record?
Field Site: This project will primarily take place at a site called Coral Gardens (or Mitchell Rocks), off the southern tip of Ambergris Caye, Belize . Coral Gardens sits between the 1,116 hectare Hol Chan Marine Reserve, created in 1987, and Caye Caulker Marine Protected Areas, but Coral Gardens has no protected status. Hol Chan is the oldest marine reserve in Belize and ‘no-take’ regulations are actively enforced within the park. The heart of Coral Gardens is composed of acroporid-dominated patches that are variably connected to one another. Five semi-permanent transect locations were established in 2012 and each transect end has been marked with rebar stakes, underwater buoys, and high resolution GPS measurements. Preliminary data from three field studies at Coral Gardens in 2011, 2012, and 2013, suggest that A. cervicornis populations (as well as A. palmata and the hybrid species A. prolifera) are anomalously healthy and robust at Coral Gardens, Belize. All three acroporid species (A. cervicornis, A. palmata, and A. prolifera) are present, live acroporid tissue abundance is high in many places, actively growing branch tips are common, and new coral recruits can be seen colonizing recently dead coral rubble and framework. Why is this isolated patch “healthier” than the surrounding acroporid-dominated patches? How do grazer dynamics, hydrology, and other physical and biological variables differ between sites? How might these different reefs look in the geologic record?
Factors potentially impacting acroporid abundance at Coral Gardens:
Resilience or persistence of robust acroporid coral growth at Coral Gardens may be a measure of how well this particular system is adapted to some of the global or regional stressors that are affecting coral reefs worldwide. Or the abundance of corals at this site could reflect unique or pristine environmental conditions off this part of Belize. Coral reefs are dynamic systems and many physical, biological, chemical, and ecological variables potentially influence coral reef community dynamics and coral population health in time and space (e.g. Kubicek et al., 2012 and references therein). Several variables that will be measured in this study that could be contributing to the overall resilience of Coral Gardens acroporids and/or the perceived spatial heterogeneity in live coral abundance, grazer populations, and A. cervicornis morphology across this reef. Students will have the opportunity to investigate and quantify the spatial and/or temporal variability in algal growth, live coral growth, herbivory and grazer dynamics, net carbonate accretion, bioerosion, genetic heterogeneity, water quality and hydrodynamics, and geochemistry of water and coral carbonate across the reef. We also hope to collect and obtain radiocarbon dates from several short cores through the reef to determine whether acroporid growth has been continuous (indicating a true refugia) or sporadic over the last several decades.
Project Aims
The aim of the work proposed here is to document a large and healthy acroporid-dominated reef system at Coral Gardens off Ambergris Caye, Belize, characterize the conditions under which acroporids are thriving, model current and potential reef accretion rates across Coral Gardens, and determine whether current live coral abundance is representative of the recent geologic record at this site. In order to do so, we will conduct a detailed observational, photographic, and quantitative habitat survey of Coral Gardens for comparison with similar data collected in 2012 and 2013 (one transect location was established in 2011 and four additional transect locations were established in 2012; and each site has been revisited since). We will particularly focus on a comparative study of two distinct areas of the Coral Gardens reef system where dramatic differences in acroporid populations exist. We will characterize differences in coral morphology, size of acroporids, percentage live tissue, branch-to-area ratio, canopy depth, fish assemblages, urchin density, temperature and light variability at each transect location, and compare with coral and zooxanthellae genetic diversity across the reef. We hope to determine whether these differences are a reflection of genetic heterogeneity in the coral hosts or algal symbionts or environmental conditions at different sites. A key goal of this study will be to determine whether acroporid growth has been persistent through the 1980’s die-off (a true refugium) or whether the corals are “coming back”. To address this we plan to collect, radiocarbon date, and analyze several cores below living coral at this site. In addition, we hope to collect geochemical data (d18O, d13C, and Sr/Ca, DIC) from water and core samples to better characterize current and past environmental conditions at Coral Gardens. The hope is that observational, photographic, and geochemical data collected from Coral Gardens acroporids will provide the information necessary to better characterize the ideal conditions for these species in the future, and interpret environmental conditions in recent geologic time. Our hope also is that these data will also be valuable to those working to improve the life expectancy for threatened coral reef systems.
Potential Student Projects
We envision a wealth of possible student research projects at this location. Depending on student interests and logistics, the project ideas below might be addressed collaboratively by individuals, or small groups of students:
- Is live A. cervicornis increasing or decreasing at Coral Gardens and is this reef in a phase transition from coral to algae dominance like most other reefs in the Caribbean? We will conduct a comparative quantitative analysis of live coral tissue, macroalgae, and bare rock abundance in 2012, 2013, and 2014 to determine whether the coral or algae are increasing or in decline within and between sites at Coral Gardens and document maximum extent of acroporid growth using GIS mapping, aerial surveys, and satellite imagery (if available) from 2012-2014. 2-4 students.
- How persistent has acroporid growth been through the well-documented period of coral decline and in recent geologic time? We plan to retrieve several short cores through recent live reef to determine whether a hiatus between new and old coral growth exists and whether this reef is persistent or in recovery. We will create a stratigraphic model and facies map for reef growth using radiocarbon dating, coral species identification, and taphonomic data from acroporids in core material. 2-3 students.
- What is the current carbonate budget for Coral Gardens and how heterogeneous is it across the site? We will determine roughly how much carbonate accretion is taking place at Coral Gardens reef by estimating live coral abundance, coral growth rates, and skeletal density of coral samples. We can estimate how fast carbonate is being excavated from reef framework via quantification of net annual bioerosion by grazers (fish and urchins) and macroborers and comparison with data from 2012 and 2013. We will place our estimates in the context of previous estimates of reef accretion (often based on non-branching coral growth) and the literature. We would also like to explore how quickly recently dead Acropora coral is incorporated into solid reef framework and detrital sediment via analysis of rates of onset of post-mortem encrustation and sponge bioerosion at the base of acroporid colonies. 2-4 students.
- Why is coral growth at this site exceptionally robust and how might designation of marine protected status at Coral Gardens impact coral abundance at this site? We will attempt to quantify the physical, biological, and environmental conditions that might make Coral Gardens an exceptional location for coral growth. We hope to compare temperature, light, and salinity data at Coral Gardens with sites across the Caribbean and we could conduct a geochemical characterization of water and coral skeletal carbonate for comparison with similar geochemical data from other acroporid corals in time (Holocene) and space (Barbados and the Dominican Republic) (Greer et al., 2009). We could also explore the degree to which coral predation (damselfish, snails, worms) and disease are impacting acroporid populations at Coral Gardens. We may also examine piscivore and herbivore trophic dynamics at Coral Gardens in comparison with the nearby Hol Chan Marine Reserve to determine whether significant spillover from the protected reef is affecting grazing effectiveness and coral recruitment at Coral Gardens. 1-3 students.
- How heterogeneous are the variables affecting live coral abundance at this site and what are the implications of heterogeneity for interpretation of the fossil record? We will explore how heterogeneous coral, algae, and herbivore populations across this reef and how variations in herbivory might be impacting live coral cover. We will compare current velocity and vector data across the reef with vector analysis of coral branches at each site to determine hydrological variation and effects on coral framework properties. We will examine morphological and/or phenotypic variability in acroporids within and between transect sights, and compare with genetic data from morphological variants collected in 2013 with the purpose of determining whether diversity of form is driven by genetic or environmental variation. 2-4 students.
 Logistics and Field Conditions
Logistics and Field Conditions
Accepted students will spend 2.5 weeks in Belize and 1.5 weeks on the Washington and Lee campus after Belize.
Working conditions and safety in the field and lab: Working in the water can be physically demanding, whether using SCUBA or a snorkel. All students on this project are required to be proficient swimmers. Students will be spending long hours on boats and in the water (up to 8 hours per day). We will almost always be in at least 10 feet of water so there will be no place to stand at the site, except for back at the boat. Most days will involve moderate wave action and mild tidal currents. Rain, cool weather on cloudy days, or windy conditions will not prevent data collection. We are working close to the tropics, and participants should be prepared to avoid sunburn and dehydration.
While a few projects are appropriate for snorkelers (primarily photo survey-based projects), most projects require SCUBA certified divers for efficient and effective data collection. Even for projects that do not explicitly require SCUBA, students will benefit from being SCUBA certified prior to the project period as SCUBA allows divers to spend more relaxed time at depth on the reef making observations and collecting data. Coral Gardens is an excellent location for newly certified divers as our maximum depth will be ~10 meters (30 feet) and the ability to look closely at the reef while diving can provide a much more informative and enjoyable reef experience. Newly certified divers will have an opportunity to develop their SCUBA and underwater research skills during the first few days at the field site. The ability to participate as a diver will also allow students to be fully involved in all aspects of data acquisition for their own project and collaboratively with their peers. Each day, students will be divided into small working groups based on shared project interests and logistical constraints. These groups may change as projects evolve and we will attempt to expose students to the widest possible variety of subject areas. SCUBA certification will enhance all student learning experiences. All students must provide their own snorkeling equipment (mask, fins, booties, snorkel, and wetsuit or equivalent) but all SCUBA gear and tanks will be provided. More information will be provided to accepted participants. If the scientific goals and methods of this project excite you, but the logistics of getting SCUBA certified before the start of the project will be challenging for you, please contact Lisa Greer.
In Belize we will be staying at the TREC Marine Field station (http://www.belizemarinetrec.com/) in comfortable but basic group accommodations. All but one to two days will likely be spent on and in the water with optional trips to Mayan Ruins, local caves, or recreational dive sites. Afternoons and evenings will be spent compiling and transcribing field data, cleaning and preparing gear for the next day, and reading and discussing relevant literature. We will meet each evening after dinner to discuss the progress of each individual and what the aims are for the next day. All students will be expected to work collaboratively and aid other field projects as needed.
NOTE: Please address your swimming proficiency and whether or not you expect to be SCUBA certified prior to the beginning of the project in your Keck application. If you have any questions about your physical fitness for the project, please contact Greer with questions.
In Lexington (Washington and Lee campus) students will stay in campus housing. While at W&L students will process samples and data (from core material to photographic data) for the post project period. Students will have access to analytical equipment such as a Scanning Electron Microscope, X-ray diffractometer, and Stable Isotope Mass Spectrometer for water, core, or sample analyses, and all photo-analysis software. Some students will return to their home with samples for petrographic, core description, or geochemical analyses and many students will leave Lexington with photographic data for further processing and analyses.
Air travel: Students will travel to Belize International Airport from their home location. To keep travel costs low, they may be asked to drive some distance to a major airport if a less-expensive alternative to local travel is available. All project participants will travel together from Belize International Airport to Ambergris Caye. At the end of fieldwork all project participants will travel together from Ambergris Caye to Washington and Lee University via Belize City and Roanoke, Virginia. At the end of the project participants will travel back to their original destinations.
Preferred student background
We are looking for students with interdisciplinary scientific interests (see below for recommended course preparation). We are also looking for students who can become comfortable collecting data in the water relatively quickly. While it is not necessary that every participant be a certified diver, all participants must be strong swimmers (see above), and many projects will require SCUBA certification at the beginning of the project.. Coursework or a demonstrated interest in some subset of the following areas is preferred; sedimentology, stratigraphy, paleontology, oceanography, marine geology, environmental studies, biology, geochemistry, or historical geology.
For more information please contact Lisa Greer (greerl@wlu.edu)
References
Aronson, R.B. and Precht, W.F., 1997, Stasis, biological disturbance, and community structure of a Holocene coral reef. Paleobiology 23:326-346.
Aronson, R.B. and Precht, W.F., 2001, White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia, 460:25-38.
Greenstein, B.J., Curran, H.A., and Pandolfi, J.M., 1998, Shifting ecological baselines and the demise of Acropora cervicornis in the Western North Atlantic and Caribbean Province: A Pleistocene perspective. Coral Reefs 17:249–261.
Greer, L., Jackson, J., Curran, H.A., Guilderson, T., and Teneva, L., 2009, How vulnerable is Acropora cervicornis to environmental change? Lessons from the early to mid-Holocene. Geology, 37:263-266.
Hughes, T.P., 1994, Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science, 265(5178):1547-1551.
Hubbard, D.K., Gladfelter, E.H., and Bythell, J.C., 1994, Comparison of biological and geological perspectives of coral-reef community structure at Buck Island, U.S. Virgin Islands, in R. N. Ginsburg, editor, Proceedings of the colloquium on global aspects of coral reefs: health, hazards and history. Rosenstiel School of Marine and Atmospheric Science, University of Miami, Miami, Florida, USA, 201-207
Jackson, J.B.C., 1992, Pleistocene perspectives on coral reef community structure. American Zoologist, 32:719-731.
Jackson, J.B.C., Kirby, M.X., Berger, W.H., Bjorndal, K.A., Botsford, L.W., Bourque, B.J., Bradbury, R.H., Cooke, R., Erlandson, J., Estes, J.A., et al., 2001, Historical overfishing and the recent collapse of coastal ecosystems. Science, 293:629–638.
Kubicek, A., Muhando, C., and Reuter, H., 2012, Simulations of Long-Term Community Dynamics in Coral Reefs – How Perturbations Shape Trajectories. PLoS Computational Biology 8(11) , art. no. e1002791.
Miller, S.L., Chiappone, M., Rutten, L.M., and Swanson, D.W., 2009, Population status of Acropora corals in the Florida Keys. Proc. of the 11th Int. Coral Reef Symp., Ft. Lauderdale, FL, 775-779.
Pandolfi, J.M., Jackson, J.B.C., Baron, N., Bradbury, R.H., Guzman, H.M., Hughes, T.P., Kappel, C.V., Micheli, F., Ogden, J.C., Possingham, H.P., and Sala, E., 2005, Are U.S. coral reefs on the slippery slope to slime? Science, 307:1725–1726, DOI: 10.1126/science.1104258.
Precht, W.F. and Aronson, R.B., 2004, Climate flickers and range shifts of reef corals, Frontiers in Ecology and the Environment, 2:307-314.
Rogers, C.S., 1985. Degradation of Caribbean and western Atlantic coral reefs and decline of associated fisheries, in Proceedings of the fifth international coral reef congress, Tahiti, C. Gabrie and M. Harmelin-Vivien, editors. Volume 6. Antenne Museum-EPHE, Moorea, French Polynesia, 491-496
Wapnick, C.M., Precht, W.F., and Aronson, R.B., 2004, Millennial-scale dynamics of staghorn coral in Discovery Bay, Jamaica. Ecology Letters, 7(4):354-361.
Williams, D.E. and Miller, M.W., 2012, Attributing mortality among drivers of population decline in Acropora palmata in the Florida Keys (USDA). Coral Reefs, 31:369-382.